
Pourquoi le rouge est sans mélange : quand le cerveau sculpte ses propres couleurs
Posez une tomate rouge sur la table, puis une mandarine à côté. Regardez-les. Le rouge vous semblera absolu, presque géométrique — un bloc sans fissure. L’orange, lui, trahit quelque chose : on y perçoit confusément du jaune et du rouge, comme un accord de piano où deux touches sonnent ensemble. Personne ne vous a appris cette différence. Personne ne vous a expliqué que certaines couleurs sont « pures » et d’autres composites. Et pourtant, vous le sentez — d’instinct, sans effort, invariablement.
Cette asymétrie est l’une des énigmes les plus tenaces de la perception visuelle. Elle a traversé un siècle et demi de neurosciences sans recevoir d’explication satisfaisante. Jusqu’en mars 2026.
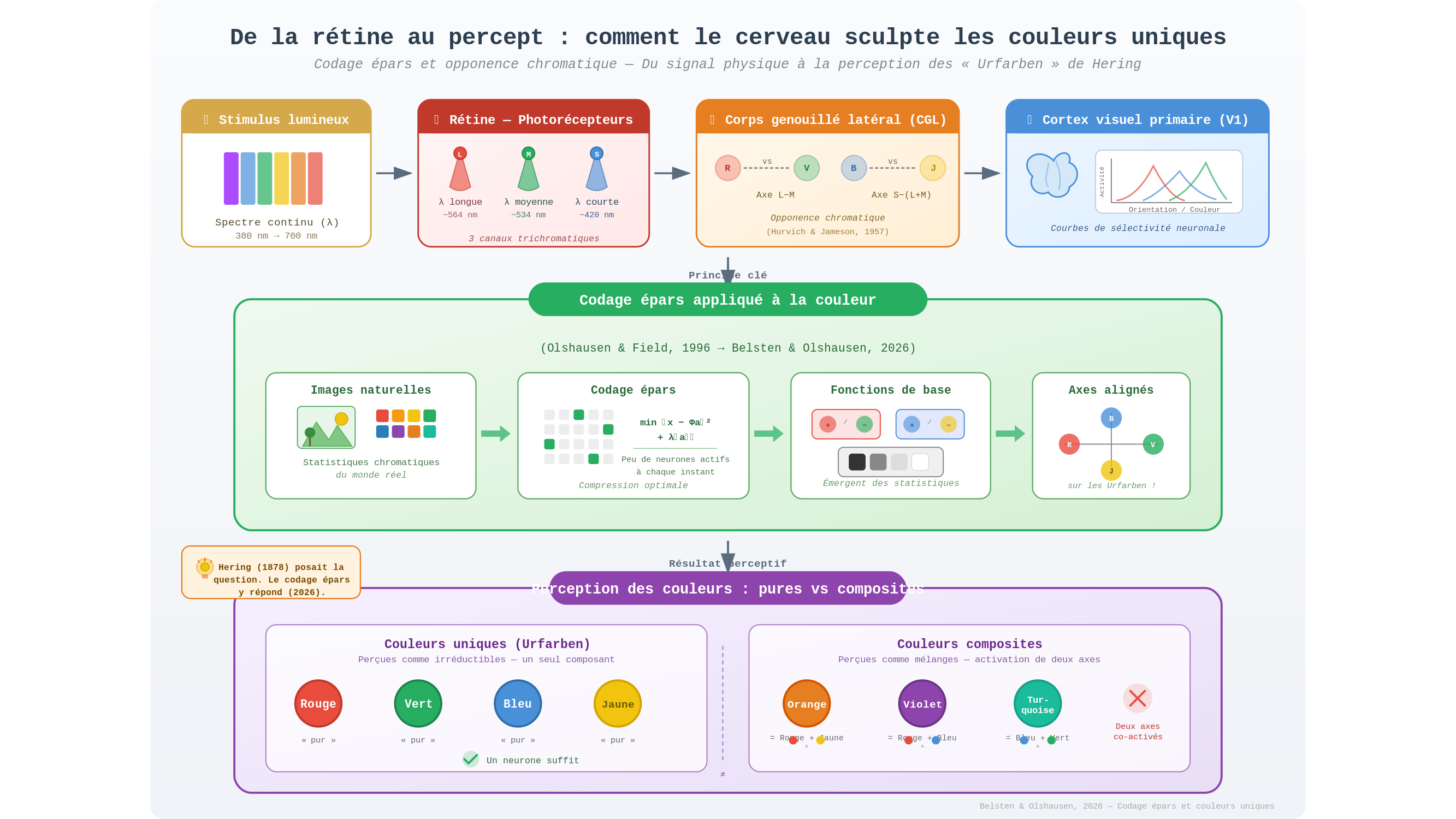
L’histoire commence en 1878, dans le laboratoire du physiologiste allemand Ewald Hering. Cet homme, que l’histoire a parfois traité un peu sèchement — il défendait des idées sur la vision héréditaire qui semblaient alors spéculatives —, décrit quelque chose d’étrange : il n’existe, pour l’œil humain, que quatre couleurs perçues comme irréductibles. Le rouge. Le vert. Le bleu. Le jaune. Toutes les autres — l’orange, le violet, le turquoise — sont perçues comme des mélanges. Ces quatre teintes particulières, Hering les appellera « couleurs uniques » (Urfarben). Elles résistent à toute décomposition perceptive. Vous ne pouvez pas voir « le rouge dans le rouge ». Vous ne pouvez pas en extraire un composant. Il est là, entier, opaque à l’analyse.
Ce qui rend l’affaire troublante, c’est que la rétine ne s’en préoccupe pas. Elle possède trois familles de photorécepteurs — les cônes L, M et S, sensibles respectivement aux longueurs d’onde longues (vers le rouge-orangé), moyennes (vert) et courtes (bleu-violet). Leurs pics de sensibilité ne coïncident pas avec les quatre couleurs uniques de Hering. La physiologie de l’œil ouvre la question sans la refermer, comme si elle s’en lavait les mains avant de la passer à quelqu’un d’autre.
Ce « quelqu’un d’autre », c’est le corps genouillé latéral — le CGL —, une structure enfouie au cœur du cerveau, premier relais du signal visuel entre la rétine et le cortex. C’est là que s’organisent les premières voies dites « d’opponence chromatique » : des circuits qui codent non pas une couleur absolue, mais une opposition — rouge contre vert, bleu contre jaune. L’idée remonte aux travaux d’Hurvich et Jameson dans les années 1950, eux-mêmes héritiers de Hering. Mais si le CGL plante le décor de l’opponence, il n’explique pas pourquoi certains points de cet espace d’opposition deviennent perceptivement singuliers. La machinerie est là, les pièces sont en place — mais le mécanisme qui cristallise les « couleurs pures » restait invisible.
C’est un détour par une autre question, posée il y a trente ans, qui va permettre de le débusquer.
En 1996, Bruno Olshausen et David Field s’intéressent à une propriété bizarre du cortex visuel primaire, cette première aire cérébrale — dite V1 — où le signal visuel est traité après le CGL. Ses neurones présentent des courbes de sélectivité (tuning curves) très précises pour les orientations et les contours. Pourquoi cette organisation-là, plutôt qu’une autre ? Olshausen et Field proposent une réponse élégante : ces courbes ne sont pas arbitraires. Elles sont le reflet statistique du monde visuel lui-même. Les contours, les bords, les surfaces orientées sont les structures les plus fréquentes dans les images naturelles. Le cortex, au fil de l’évolution et du développement, s’est calé sur ces régularités. Il les représente avec un minimum de neurones actifs à chaque instant — c’est le « codage épars » (sparse coding). Pensez à un archiviste qui, au lieu de conserver chaque document intégralement, n’enregistrerait que les éléments qui reviennent le plus souvent dans tous ses textes : il compresse, il distille, il garde l’essentiel.
Trente ans plus tard, Alexander Belsten et Bruno Olshausen (Redwood Center for Theoretical Neuroscience, Université de Californie à Berkeley) et E. Paxon Frady (Intel Neuromorphic Computing Lab, Santa Clara) appliquent ce même raisonnement aux couleurs. L’étude, mise en ligne en mars 2026 (arXiv:2603.24293), part de 503 images naturelles calibrées spectralement — des photographies du monde réel, avec une lumière réelle et des surfaces réelles, non truquées. À partir de ces images, ils calculent les activations de cônes correspondantes et observent la distribution des couleurs naturelles dans l’espace chromatique.
Ce qu’ils trouvent est à la fois simple et vertigineux. La distribution n’est pas uniforme. Elle présente des « queues lourdes » — des excès statistiques de couleurs très saturées — dans des directions précises de l’espace des cônes. Et ces directions, calculées sans aucune hypothèse préalable sur la perception humaine, coïncident avec les quatre couleurs uniques de Hering. Le rouge. Le vert. Le bleu. Le jaune.
En d’autres termes : les couleurs que le cerveau perçoit comme « pures » sont celles que la nature produit le plus souvent, dans les cas extrêmes. Elles ne sont pas arbitraires. Elles sont les coins saillants d’une géographie statistique du monde visible. Si le cortex visuel devait coder les couleurs de façon éparse — en n’activant qu’un minimum de détecteurs pour chaque scène — ses détecteurs optimaux convergeraient naturellement vers ces quatre directions. La pureté perceptive du rouge serait alors une propriété émergente : non pas une vérité physique sur la lumière, mais le résultat d’une adaptation du système visuel à la distribution des couleurs dans son environnement.
Mais l’histoire mérite d’être lue avec un œil critique. Les auteurs sont prudents sur un point que la vulgarisation a parfois tendance à escamoter : le lien entre la géométrie de l’espace chromatique optimal et l’expérience subjective de la « pureté » reste un postulat, pas une démonstration. On parle ici du « problème de liaison » (binding problem) — cette question, toujours ouverte, de savoir comment des activités neuronales distribuées donnent naissance à une perception unifiée et qualitative. Que le modèle de codage épars produise des détecteurs alignés sur les couleurs de Hering est un résultat robuste. Que ces détecteurs soient la cause de la singularité perceptive du rouge reste une hypothèse séduisante, pas encore une preuve.
Il faut aussi noter que l’étude repose sur 503 images — un corpus soigneusement calibré, mais qui représente une tranche culturellement et géographiquement située du monde visuel humain. Les couleurs dominantes dans une forêt équatoriale, une toundra arctique ou un désert de sable ne sont pas les mêmes. Est-ce que des primates vivant dans des environnements radicalement différents percevraient des « couleurs uniques » différentes ? La question n’est pas rhétorique : certaines espèces possèdent quatre ou cinq types de cônes — les squilles (mantis shrimps), ces crustacés aux yeux extraordinairement complexes, en auraient jusqu’à seize. Leur monde chromatique, s’il obéit aux mêmes principes de codage épars, devrait présenter une tout autre géographie de couleurs « pures ». Ce serait une façon de tester l’hypothèse, et les auteurs l’évoquent eux-mêmes comme piste.
Pendant cent quarante-huit ans, la question de Hering est restée sans réponse parce qu’on la cherchait au mauvais endroit — dans la physique de la lumière, dans l’anatomie de l’œil, dans la chimie des photorécepteurs. La réponse, si elle se confirme, vient d’ailleurs : du monde lui-même, de sa texture statistique, de la façon dont la nature distribue les couleurs dans l’espace et dans le temps. Le rouge serait pur non pas parce qu’il est simple, mais parce que le cerveau s’est calibré sur un monde où il arrive souvent, intensément, sans mélange.
Ce qui laisse une question en suspens, presque vertigineuse : si nos ancêtres avaient évolué dans un monde aux couleurs différentes — plus d’ultraviolet, moins de vert, des surfaces aux spectres de réflexion redistribués —, est-ce que le rouge existerait encore ? Est-ce que nos couleurs pures sont les nôtres, ou celles de la Terre ?
Figures originales du paper






Sources
Belsten A., Frady E. P., Olshausen B. A. — Unique hues from sparse coding of natural color statistics — arXiv:2603.24293 (mars 2026)
Olshausen B. A. & Field D. J. — Emergence of simple-cell receptive field properties by learning a sparse code for natural images — Nature, 381, 607–609 (1996). DOI: 10.1038/381607a0


