
Il y a, au cœur de la théorie classique du comportement de chasse, une hypothèse si bien enfouie qu’on la remarque à peine : le mouvement appelle le mouvement. Le prédateur répond au déplacement de la proie ; la fuite déclenche la poursuite. Ce couplage paraît si naturel, si fondé dans l’observation du vivant, qu’on en a fait presque un axiome. Que se passerait-il si cet axiome était moins solide qu’on ne le suppose ?
C’est la question qu’explore l’écologue comportementaliste Inon Scharf dans une étude publiée en 2025 dans les Annals of the New York Academy of Sciences. Sa méthode est digne d’intérêt — et mérite d’être présentée pour ce qu’elle est, sans l’enrober de naturalisme hâtif : Scharf n’a pas observé des araignées ni filmé des prédateurs dans leur habitat. Il a construit un monde entièrement numérique, peuplé d’agents simulés dont les comportements obéissent à des règles explicitement codées. Ce n’est pas une étude sur des animaux ; c’est une démonstration par le calcul. La distinction n’est pas de détail.
Le cadre formel qu’il mobilise s’appelle un modèle individu-centré — en anglais, individual-based model ou IBM. L’idée est de simuler chaque entité séparément, avec ses propres règles de déplacement, plutôt que de décrire le système par des équations globales. Ce qui en émerge, ce sont des comportements collectifs que personne n’a explicitement programmés — un peu comme des embouteillages qui naissent de règles de conduite individuelles pourtant banales. Chaque agent prédateur de Scharf obéit à des règles simples : il se déplace, il rencontre des proies, il les capture selon des probabilités définies. Les proies, elles, sont soit immobiles, soit dotées d’une certaine vitesse de déplacement. En faisant varier trois paramètres — la distribution spatiale des proies, la directionnalité des trajectoires et la vitesse des proies mobiles — Scharf obtient un résultat qui dérange l’intuition dominante.
Dans certaines configurations, les agents prédateurs actifs — c’est-à-dire ceux dont les règles de déplacement sont codées pour la poursuite — produisent des taux de capture plus élevés sur des proies immobiles que sur des proies mobiles. Les règles de déplacement conçues pour la chasse génèrent, dans ces configurations précises, une efficacité inattendue contre des cibles qui ne bougent pas. Ce résultat n’a pas été programmé : il émerge du modèle. C’est cela, en réalité, qui mérite attention.
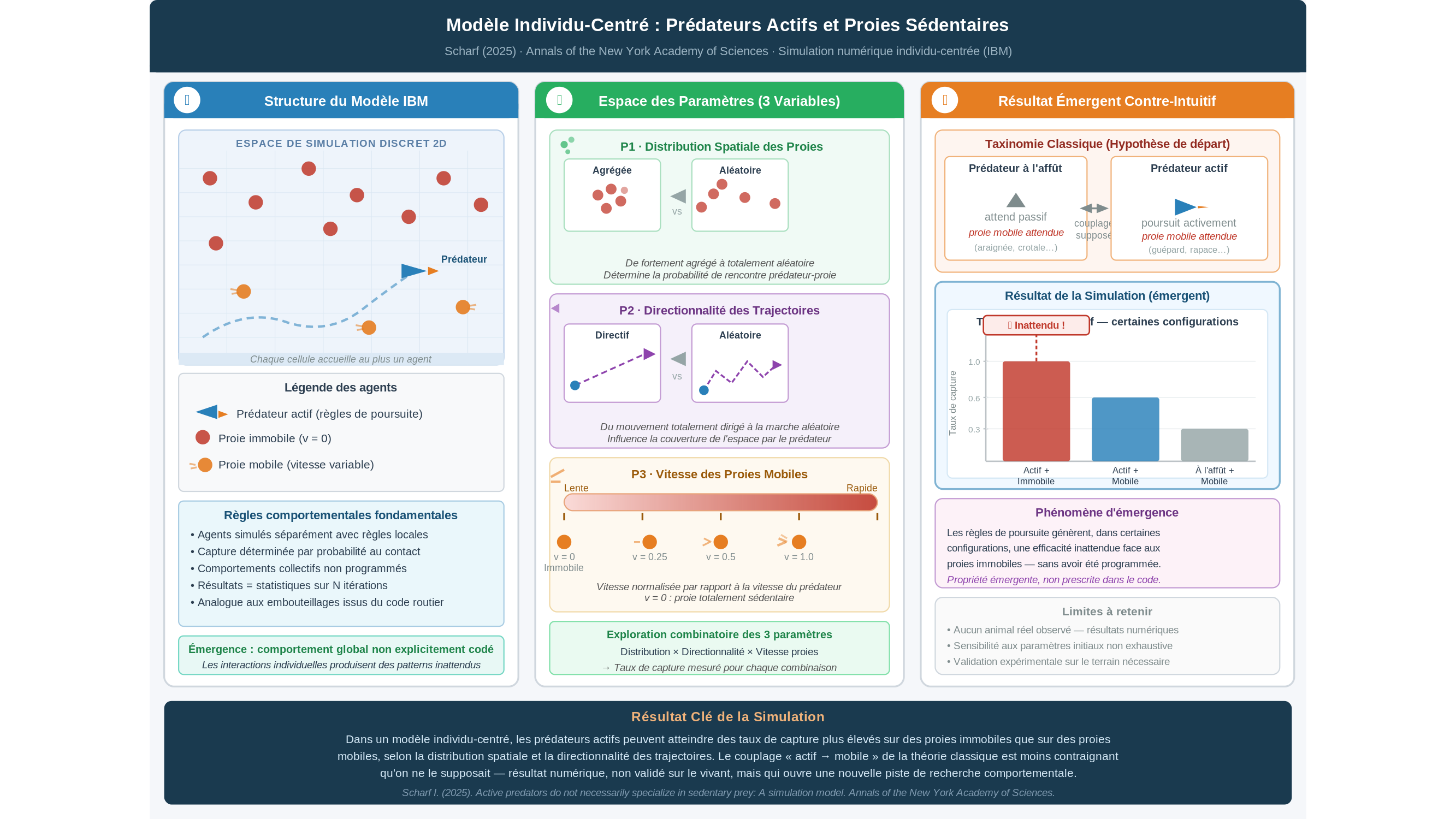
La taxinomie classique de l’écologie comportementale oppose deux grandes stratégies : les prédateurs à l’affût — l’araignée immobile dans sa toile, le crotale tapi entre deux pierres, qui attendent que la proie vienne à eux — et les prédateurs actifs, qui parcourent l’espace, répondent aux signaux visuels ou olfactifs et adaptent leurs trajectoires. Cette bipartition, commode, repose sur un postulat implicite : chaque type de chasseur est optimisé pour un type de proie. Les actifs chassent du mobile ; les sédentaires attendent du passant. La simulation de Scharf met en évidence que ce couplage supposé est peut-être moins contraignant qu’on ne le pensait — du moins dans l’espace abstrait d’un modèle numérique, et c’est précisément là que la prudence doit entrer en scène.
Car il y a une limite que Scharf lui-même ne cherche pas à dissimuler : ses résultats restent entièrement dans l’ordre du calculable, pas du biologique. Aucun animal réel n’a été observé, aucun comportement n’a été mesuré sur le terrain. On peut aussi s’interroger sur la robustesse du modèle lui-même : un IBM est structurellement sensible aux hypothèses de départ — règles de déplacement, densité des agents, géométrie de l’espace simulé. Des paramètres légèrement différents produiraient-ils les mêmes résultats ? L’article ne le dit pas, et c’est une limite structurelle de tout modèle individu-centré. Passer du modèle au vivant exige en outre une validation expérimentale que cette étude ne fournit pas. Ce n’est pas un reproche — un modèle bien construit peut produire des hypothèses fécondes que l’expérience se chargera de confirmer ou d’infirmer. Mais c’est une frontière qu’il faut nommer.
Ces résultats invitent à réfléchir à la thanatose — stratégie évolutive par laquelle certaines proies simulent leur propre mort pour échapper à un prédateur, dont le possum virginien est l’exemple canonique. Si des prédateurs actifs se révèlent capables de capturer efficacement des proies immobiles, cela pourrait, dans une lecture évolutive, nuancer l’idée que la thanatose constitue une défense universellement efficace contre tout type de chasseur. Mais il convient de souligner que cette inférence demeure conjecturale : le modèle de Scharf ne modélise pas la thanatose, et la pression de sélection est un niveau d’explication que la simulation ne peut atteindre seule.
Ce qui demeure, une fois les précautions prises, c’est une invitation à réexaminer une catégorie. Quand on dit qu’un prédateur est « actif » ou « à l’affût », on croit décrire une propriété de l’animal. Peut-être décrit-on aussi une relation — une relation à un type de proie, dans un environnement particulier, sous des contraintes spécifiques. La biologie du comportement est riche en catégories qui semblent naturelles jusqu’au moment où un modèle, justement parce qu’il est abstrait, met en évidence que leur contenu était plus contingent qu’absolu. C’est là, peut-être, l’un des usages les plus précieux de la simulation : non pas imiter la nature, mais révéler les présupposés que nous projetons sur elle.
Que se passerait-il si la distinction entre prédateur actif et prédateur passif était moins une propriété de l’organisme qu’un artefact de notre façon de découper le monde ? Ce n’est pas Scharf qui pose cette question — son article reste soigneusement dans les limites de son modèle. Mais c’est la question que le modèle laisse ouverte, comme une ombre portée sur la certitude ordinaire.
Sources
Scharf I. (2025), « Active predators do not necessarily specialize in sedentary prey: A simulation model », Annals of the New York Academy of Sciences. PMID : 40400223. DOI : 10.1111/nyas.15379 (à confirmer — abstract seul disponible au stade de rédaction).
Huey R.B. & Pianka E.R. (1981), « Ecological consequences of foraging mode », Ecology, 62(4), 991–999. DOI : 10.2307/1936998 — référence classique sur la dichotomie affût/poursuite dans les stratégies de prédation.


