
Trois heures d’affilée. C’est le temps pendant lequel un mâle rorqual à bosse peut chanter sans interruption, suspendu entre deux eaux dans l’obscurité des zones tropicales, tête inclinée, nageoires légèrement écartées. La séquence qu’il produit obéit à une architecture précise : thèmes, phrases, unités sonores emboîtées les unes dans les autres selon un ordre hiérarchique qui n’est pas sans évoquer celui d’une composition musicale. Ce qui distingue ces productions vocales — non pas dans l’absolu, car les oiseaux chanteurs présentent sur certains critères une complexité vocale comparable, parfois supérieure, qu’il s’agisse de la grive polyglotte ou de l’oiseau-lyre australien — c’est ailleurs. C’est la durée exceptionnelle des séquences, leur organisation simultanée sur plusieurs niveaux syntaxiques, et surtout la dynamique singulière par laquelle un répertoire entier se propage et se transforme à l’échelle d’une population entière, d’une saison à l’autre, d’un individu à l’autre, d’un océan à l’autre. Rien de tel n’a été documenté chez aucune autre espèce pour ce phénomène précis de transmission et d’évolution culturelle collective.
Depuis que Roger Payne et Scott McVay ont publié, en 1971, la première description rigoureuse de ces chants — une étude parue dans Science qui a littéralement redessiné le regard porté sur les cétacés —, les biologistes n’ont cessé d’enregistrer, de cartographier, de modéliser ces séquences vocales. Ce qui captivait surtout les chercheurs, c’était la dimension collective du phénomène : comment une phrase nouvelle se glisse dans le répertoire commun, comment les mâles l’adoptent, comment une innovation acoustique traverse en quelques mois le Pacifique tout entier. C’est de la biologie culturelle, au sens presque plein du terme.
Ce que personne ou presque n’avait regardé de près, en revanche, c’est ce qui se passe entre deux chants consécutifs du même individu. Dans l’espace de quelques minutes, entre deux plongées, pendant que l’animal reprend son souffle en surface — que se passe-t-il dans la production vocale ? Y a-t-il une continuité, une dérive, un ajustement ? La question peut sembler secondaire, microscopique à l’échelle d’une littérature construite sur des dizaines d’années d’observations. Elle ne l’est peut-être pas.
C’est précisément là que porte une étude d’Eduardo Mercado, Monika Ashour et Sara McAllister, publiée en 2022. Les auteurs y formulent une hypothèse qui oblige à revisiter plusieurs décennies d’interprétations : la variabilité observée entre les chants successifs d’un même mâle ne serait pas le produit d’une dérive aléatoire, ni d’un programme vocal imparfaitement stable, mais le signe d’une régulation active. Ils appellent ce mécanisme supposé le « contrôle cognitif » de la production vocale — formulation qui mérite qu’on s’y arrête un instant.
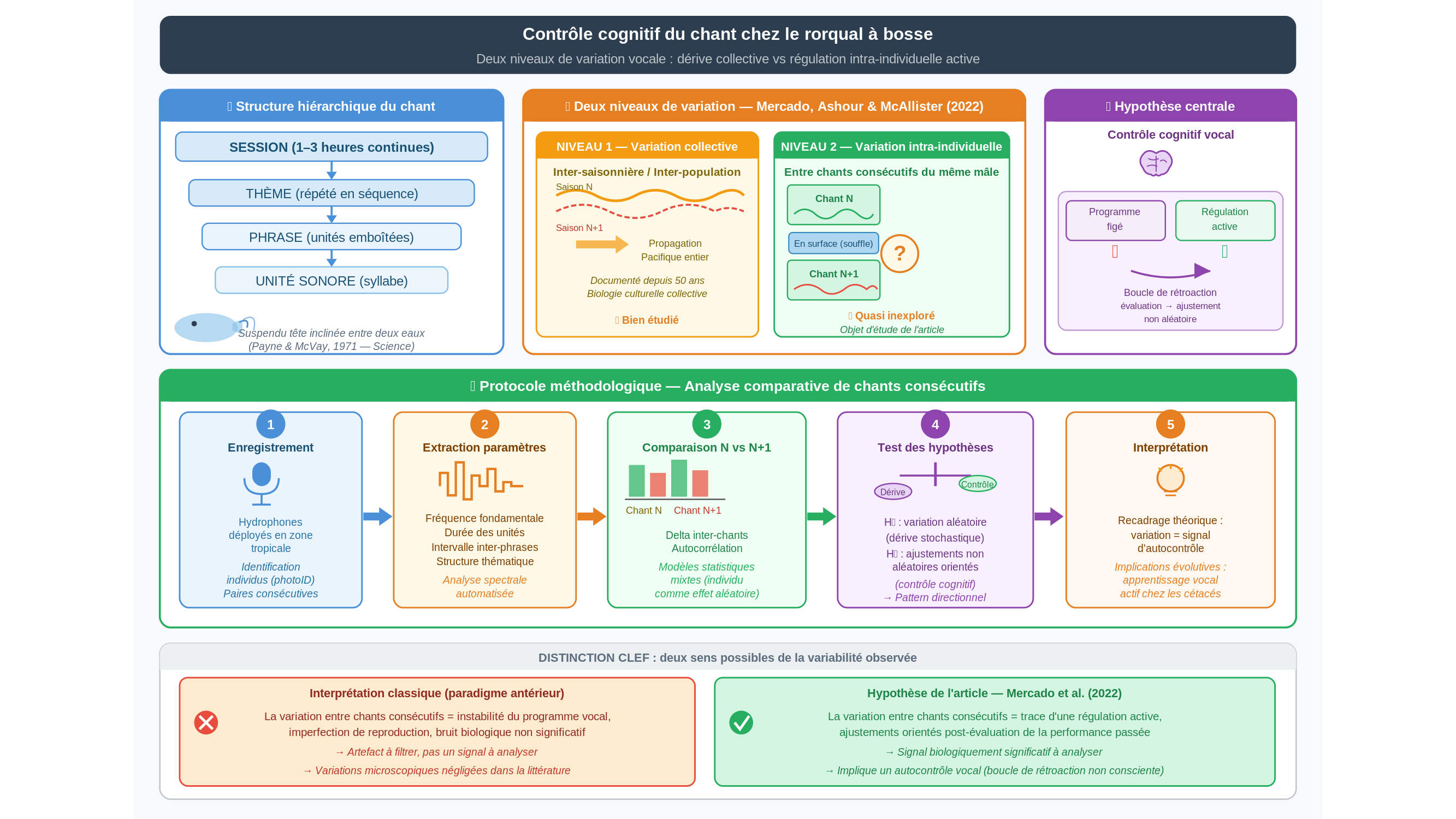
Le mot « cognitif » est ici technique au sens strict. Il ne signifie pas que l’animal réfléchit au sens où nous l’entendons ordinairement. Il désigne quelque chose de plus précis et de plus circonscrit : la possibilité que le rorqual module sa séquence sonore en s’appuyant sur une évaluation de sa propre performance passée, plutôt qu’en rejouant mécaniquement un modèle figé. Ce serait une boucle de rétroaction — une forme d’autocontrôle vocal, pas nécessairement conscient, mais orienté, au sens où il produirait des ajustements non aléatoires. La différence, en somme, entre suivre un programme et surveiller activement sa propre exécution.
Pour saisir en quoi ce changement de regard importe, considérons la distinction que les auteurs introduisent entre deux niveaux de variation. Le premier est bien documenté : c’est la variation collective, inter-saisonnière, le glissement progressif du répertoire d’une population entière d’une année sur l’autre. Ce niveau a été étudié avec soin depuis Payne et McVay. Le second est bien plus fin : c’est la variation intra-individuelle, les différences que l’on observe entre deux sessions vocales successives du même animal. Selon Mercado et ses collègues, ce niveau a été systématiquement sous-estimé — confondu avec du bruit résiduel, attribué à des fluctuations mineures sans conséquences — alors qu’il pourrait être porteur d’une information cruciale sur les capacités de traitement vocal de l’animal.
Une analogie peut éclairer la différence. Supposons qu’on cherche à comprendre comment un musicien de jazz joue le même standard soir après soir. On peut étudier la façon dont ce standard a évolué sur cinquante ans, tracer la propagation des innovations stylistiques d’une époque à l’autre, comparer les versions de différentes générations. C’est de l’histoire musicale — légitime, instructive. Mais elle ne dit rien sur ce que le musicien fait d’une mesure à l’autre : l’écoute de sa propre phrase, les légères corrections de tempo, le choix de relancer ou de conclure. Mercado et ses collègues soutiennent que la recherche sur le chant des rorquals s’est longtemps cantonnée au premier type d’analyse, et qu’il est temps de conduire le second.
Il faut ici prendre la mesure de la limite méthodologique, parce qu’elle est sérieuse. L’argument des auteurs reste pour l’essentiel une reformulation des cadres d’interprétation disponibles — un appel à changer de focale — plus qu’une démonstration expérimentale robuste d’un mécanisme de contrôle cognitif clairement identifié. L’étude ne précise pas non plus le corpus exact sur lequel repose l’analyse : nombre d’individus suivis, volume d’enregistrements, méthode d’analyse acoustique. Pour évaluer la solidité de l’hypothèse, ces détails comptent. La question de ce que « contrôle cognitif » signifie exactement pour un mammifère marin non humain demeure ouverte, et la distinction entre variation active et variation passive n’est pas, sur la seule base d’enregistrements acoustiques, toujours aisée à établir rigoureusement. Les auteurs en sont conscients, et leur étude se lit autant comme un programme de recherche que comme un résultat clos — une invitation à instruire un dossier laissé en friche, plutôt qu’une réponse ferme.
Ce qui n’enlève rien à l’intérêt de la démarche. Montrer que des décennies d’observations ont peut-être mesuré le bon phénomène à la mauvaise échelle est en soi un apport. Si l’hypothèse résistait à des protocoles conçus pour la mettre directement à l’épreuve, elle aurait des conséquences pour plusieurs questions connexes : la flexibilité comportementale des cétacés, bien sûr, mais aussi les théories sur l’évolution du langage et de la musique — deux domaines où le chant des rorquals est régulièrement convoqué comme comparateur non humain. Séparer ce qui relève de la culture vocale collective et ce qui relève d’une capacité individuelle à moduler une production sonore en cours d’exécution n’est pas une distinction neutre dans ce débat.
Que le rorqual à bosse exerce sur son chant quelque chose qu’on puisse appeler un contrôle — ou qu’il en soit simplement le site, l’instrument, sans que le mot « contrôle » ajoute quoi que ce soit au réel — ce débat n’est pas purement académique. Il touche à une question bien plus ancienne : jusqu’où peut-on parler de productions vocales flexibles, ajustées, sensibles à leurs propres effets, sans que le concept se dilue en perdant ce qui le rendait opératoire ? Les cétacés, depuis quelques décennies, n’ont cessé de repousser les frontières de cette question. Nul ne sait encore si, cette fois, ils les déplacent à nouveau ou si c’est seulement notre façon de les regarder qui change.
Sources
- Payne R.S. & McVay S. (1971), Songs of Humpback Whales, Science, vol. 173, n° 3997, pp. 585-597.
- Mercado E., Ashour M. & McAllister S. (2022), Cognitive control of song production by humpback whales, PMID 36058997 (journal non accessible sans accès intégral).


