
Berlin, aube. Un merle perché sur une antenne émet son chant depuis une hauteur de fréquence inhabituellement élevée — plus aiguë que celle de ses cousins des forêts proches. En 2003, Hans Slabbekoorn et Ardie Peet ont établi que les merles urbains chantaient à des fréquences plus hautes que les individus forestiers d’une même région — probablement pour que leur signal émerge du fond sonore grave des moteurs et des rues. Ce phénomène précis ne figure pas dans la synthèse que Pavol Mikula et ses collègues ont publiée en 2025 — il sert ici d’entrée en matière, et il faut le dire clairement. Mais il incarne exactement ce que cette synthèse cherche à cerner : la ville ne supprime pas seulement des espèces. Elle les transforme. Et la transformation suit, partout, le même chemin.
Ce chemin a un nom — « homogénéisation comportementale » — et ce terme mérite qu’on s’y arrête. « Homogénéisation » : du grec homos, semblable, et génos, nature, origine. Le mot désigne en physique le processus par lequel un mélange devient uniforme. En biologie, il désigne quelque chose de plus troublant : la convergence, sous pression humaine, des manières d’agir entre individus d’une même espèce, entre populations géographiquement distantes, parfois entre espèces entières. La biodiversité ne se perd pas seulement par extinction — cette vérité-là, les sciences de la conservation l’ont en partie intégrée. Elle se perd aussi par ressemblance.
On connaissait l’homogénéisation biotique — ce phénomène par lequel pigeons, rats et moineaux domestiques colonisent toutes les métropoles au détriment des faunes locales. Voici sa contrepartie moins visible, que la synthèse de Mikula et al. propose de nommer et de cartographier : même quand l’espèce survit, même quand elle prospère numériquement, ses individus tendent à devenir comportementalement identiques. Non plus la même espèce partout — le même individu partout.
La synthèse recense dix domaines où ce nivellement opère : les réponses à la peur, les régimes alimentaires, les chants, les rythmes d’activité, les structures sociales, la propension à l’exploration, les routes migratoires, les choix de sites de reproduction… La ville n’efface pas que des espèces. Elle efface des façons d’être.
Prenons la peur — cette distance que maintient naturellement un animal face à une menace. En milieu urbanisé, les individus qui s’enfuient trop tôt ratent des ressources alimentaires concentrées ; ceux qui restent trop longtemps périssent sous la roue d’un véhicule ou la griffe d’un chat errant. Il subsiste alors, selon le cadre proposé par les auteurs de la synthèse, une tranche étroite de tolérance, et tous les survivants y convergent. C’est l’hypothèse centrale du modèle — non une démonstration expérimentale directe population par population, mais un cadre interprétatif qui s’ajuste bien aux données disponibles. Ce que ce modèle pointe n’est pas de l’adaptation au sens plein du terme : c’est un filtre qui élimine les extrêmes.
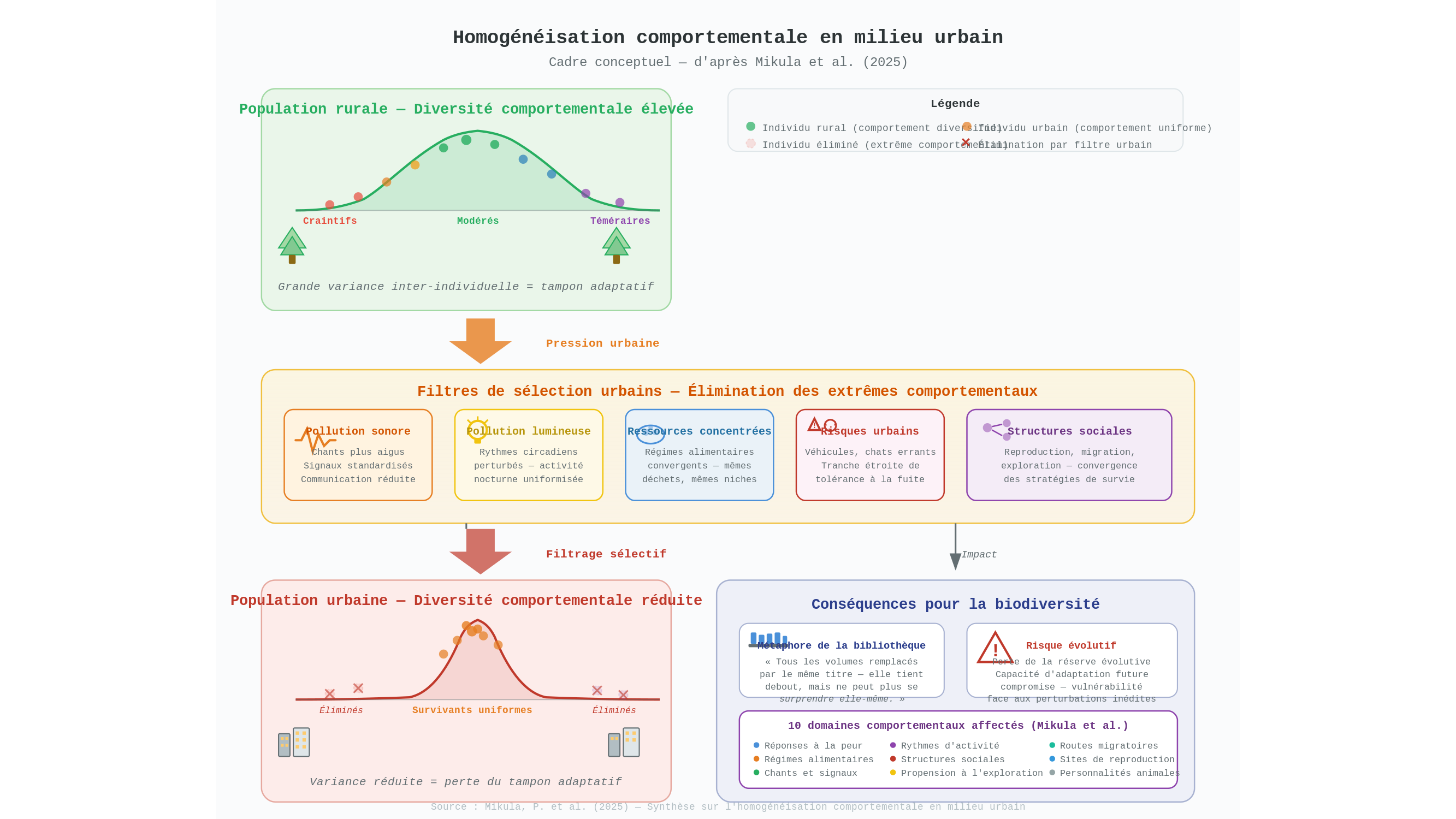
Or ces extrêmes sont précieux — et c’est là que la question devient véritablement philosophique. La diversité comportementale entre individus — les « explorateurs », les « prudents », les « téméraires » — fonctionne comme une réserve évolutive, ce que les biologistes nomment parfois un tampon adaptatif. Face à un environnement inédit, c’est souvent dans ces marges qu’une population trouve ses ressources. Une population comportementalement uniforme ressemble à une bibliothèque dont tous les volumes auraient été remplacés par le même titre : elle tient debout, elle est consultable, mais elle ne peut plus se surprendre elle-même. Et une bibliothèque qui ne peut plus se surprendre ne sait pas à quelle question elle répondra demain.
Les rythmes biologiques illustrent le même mécanisme, mais à une échelle temporelle différente. La pollution lumineuse — l’excès de lumière artificielle nocturne, ce fond de clarté permanent sous lequel vivent désormais la plupart des animaux urbains — recalibre les horloges internes des oiseaux, des insectes, des mammifères vers un profil unique. Il est difficile de ne pas voir dans cette uniformisation des rythmes une forme d’effacement plus subtile encore : non plus la diversité des corps, ni même des comportements, mais la diversité des temporalités internes. Le vivant a mis des millions d’années à diversifier ses façons de découper le temps. Nos nuits illuminées en font quelque chose de beaucoup plus simple.
Il faut pourtant nuancer. La synthèse de Mikula et al. est une proposition de cadre conceptuel, une invitation à regrouper sous un même terme des phénomènes documentés séparément dans des disciplines distinctes. Elle ne prétend pas, domaine par domaine, avoir démontré la généralité du mécanisme avec une rigueur équivalente partout. Certains des dix champs recensés reposent sur des corpus empiriques solides — les ajustements vocaux chez les oiseaux sont mesurés et répliqués depuis deux décennies. D’autres — comme l’homogénéisation des routes migratoires — demeurent moins documentés, leur mise en évidence étant techniquement difficile à l’échelle des populations entières. Il serait prématuré de lire cette synthèse comme un bilan général consolidé : elle est, plutôt, un programme de recherche — ce qui n’est pas rien, mais ce qui n’est pas non plus une démonstration.
Le paradoxe est que les instruments habituels de la conservation ne voient pas cela. Leurs métriques mesurent le nombre d’espèces — inventaires taxonomiques, listes rouges, courbes d’extinction. La dimension comportementale n’a pas encore son tableau de bord. Une espèce dont tous les individus se comportent de manière identique peut figurer en bonne santé dans les registres officiels, tout en ayant perdu une part irremplaçable de ce qui faisait sa plasticité — sa capacité à répondre à des pressions imprévues. C’est un angle mort de la biologie de la conservation, et la synthèse de Mikula et al. a le mérite de le nommer clairement.
Que faire de ce constat ? Les auteurs évoquent la nécessité d’intégrer les indicateurs comportementaux dans les évaluations de biodiversité. Mais mesurer la diversité comportementale à l’échelle d’une population — quantifier la variance des caractères individuels sur des terrains où l’on peine déjà à recenser les espèces — est une entreprise méthodologiquement redoutable. Les outils existent, pour certains taxons, dans certains milieux. Ils n’existent pas pour l’essentiel du vivant. Ce que la ville produit en matière d’effacement, elle le produit donc dans un relatif silence scientifique, dans l’écart entre ce qu’un merle chante ce matin à Berlin et ce qu’il aurait chanté si la ville n’existait pas.
Cette dernière différence — entre le chant réel et le chant possible — est peut-être la définition la plus exacte de ce que nous perdons. Non pas ce qui a disparu, mais ce qui n’aura jamais eu lieu.
À lire aussi sur Mémorabilité :
Sources
— Mikula P. et al. (2025). Behavioural homogenization as a component of human-induced biodiversity loss. PMID : 41770723. (DOI non disponible au moment de la publication — URL PubMed à vérifier avant republication.)
— Slabbekoorn H. & Peet M. (2003). Birds sing at a higher pitch in urban noise. Nature, 424, 267. DOI : 10.1038/424267a. (Cité comme illustration du phénomène d’ajustement vocal urbain ; cette étude est indépendante de la synthèse de Mikula et al.)


