
Dans les pas d’un sanglier berlinois, ce que la géométrie d’un trajet révèle
Trois heures du matin, quelque part dans Berlin. Un sanglier franchit une avenue, s’arrête, repart dans une direction que rien, en apparence, ne justifie. Son collier GPS enregistre tout — latitude, longitude, seconde après seconde. Mais entre ce nuage de points et la logique du trajet, il y a toute la distance qui sépare une partition de la musique : les notes sont là, le sens reste ailleurs.
Depuis un demi-siècle, les écologues ont appris à lire ces déplacements. D’abord à la radio, un bip dans les écouteurs signalant qu’un cerf venait de traverser telle clairière. Puis par satellite, avec des colliers GPS capables d’enregistrer la position d’un animal toutes les cinq minutes pendant plusieurs années. Les données ont explosé. La question, elle, est restée la même : pourquoi ici, et pas là ?
La méthode de référence pour y répondre porte un nom un peu aride : la fonction de sélection de pas (step selection function, ou SSF). Le principe est élégant. Pour chaque trajet réel observé, on génère une série de trajets fictifs, tirés au sort dans les mêmes conditions. Le modèle statistique ajuste ses paramètres pour maximiser la distinction entre les vrais et les inventés — et ce faisant, le modèle fait émerger les préférences inscrites dans la géométrie du mouvement. Non pas ce que l’animal dit (il ne dit rien), mais ce que la forme même de ses trajets révèle : la tendance d’un cerf à longer les lisières plutôt que les routes, d’un lynx à contourner les zones urbaines.
Ce n’est pas de la psychologie animale. C’est de la géométrie lue à rebours.
Le problème, c’est que cette méthode repose depuis ses origines sur un modèle linéaire généralisé. Autrement dit : elle suppose que chaque variable environnementale agit seule, indépendamment des autres. Or l’eau est faite d’hydrogène et d’oxygène — deux gaz qui ne mouillent pas. Ce qui compte naît parfois de la rencontre, pas des composants pris séparément. Un renard peut fuir les routes, mais seulement à proximité des habitations : cette interaction, le modèle classique ne peut pas la capturer. Il lui faudrait qu’on la lui désigne à l’avance.
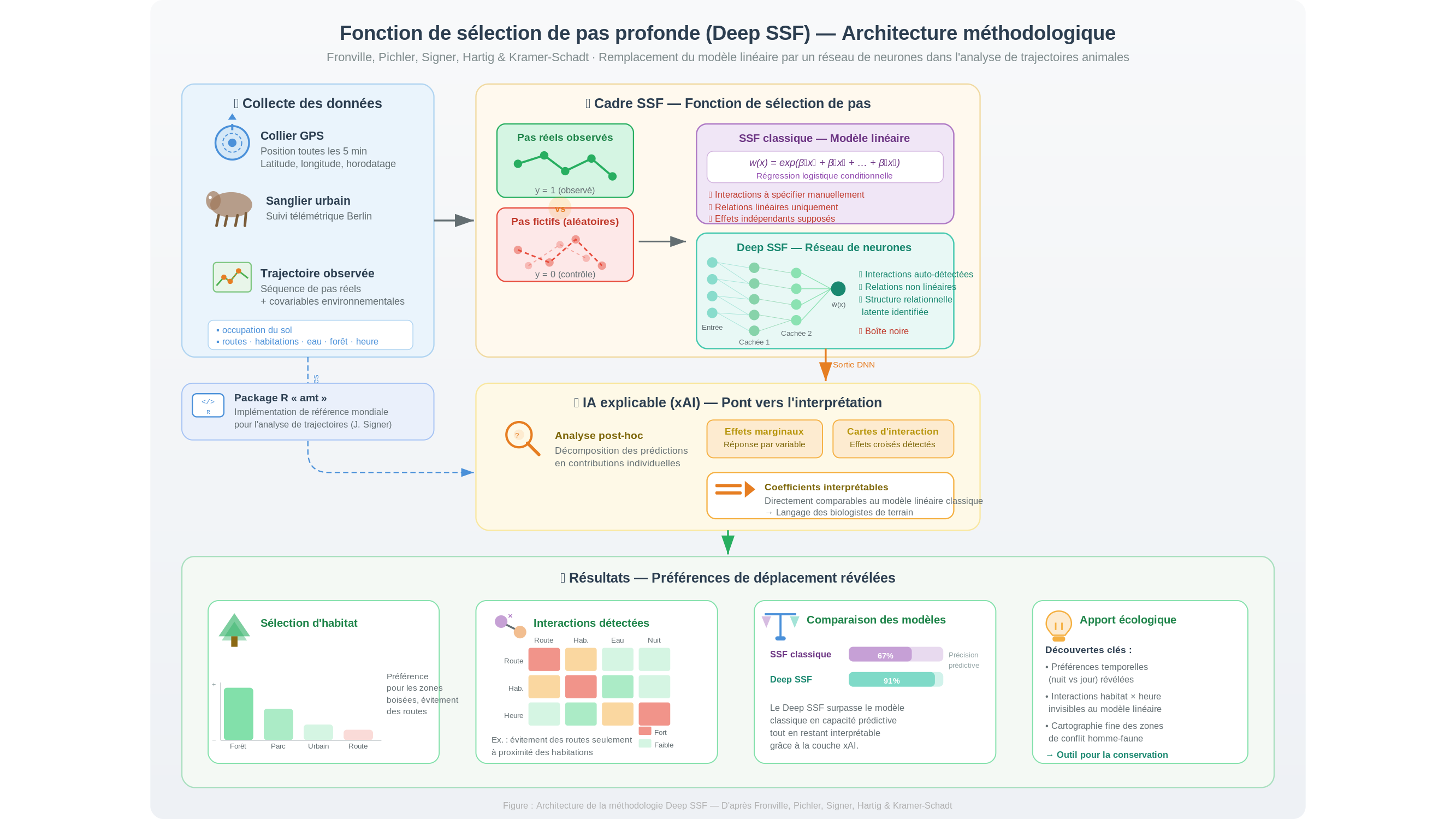
C’est précisément là que l’équipe entre en scène. Fronville, Pichler et Johannes Signer — ce dernier est aussi le développeur du package amt, référence mondiale pour l’analyse de trajectoires animales — ont uni leurs forces à celles de Florian Hartig (Université de Ratisbonne) et de Stephanie Kramer-Schadt (Institut Leibniz de zoologie et médecine de la faune sauvage, IZW, et Université Technique de Berlin). Leur idée : remplacer le modèle linéaire par un réseau de neurones profond, entraîné selon les principes de l’apprentissage profond (deep learning), et adossé à l’architecture même des SSF existantes.
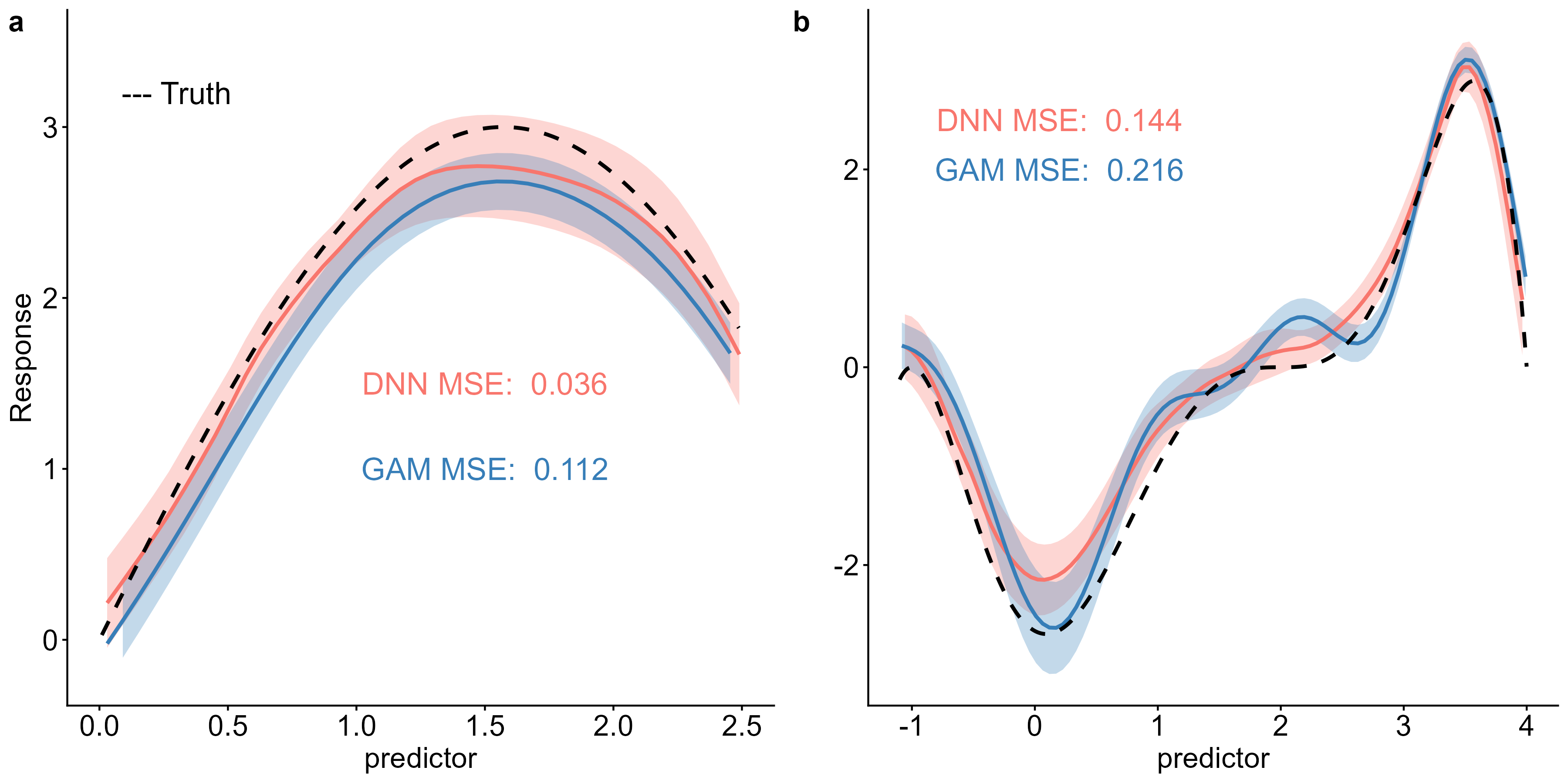
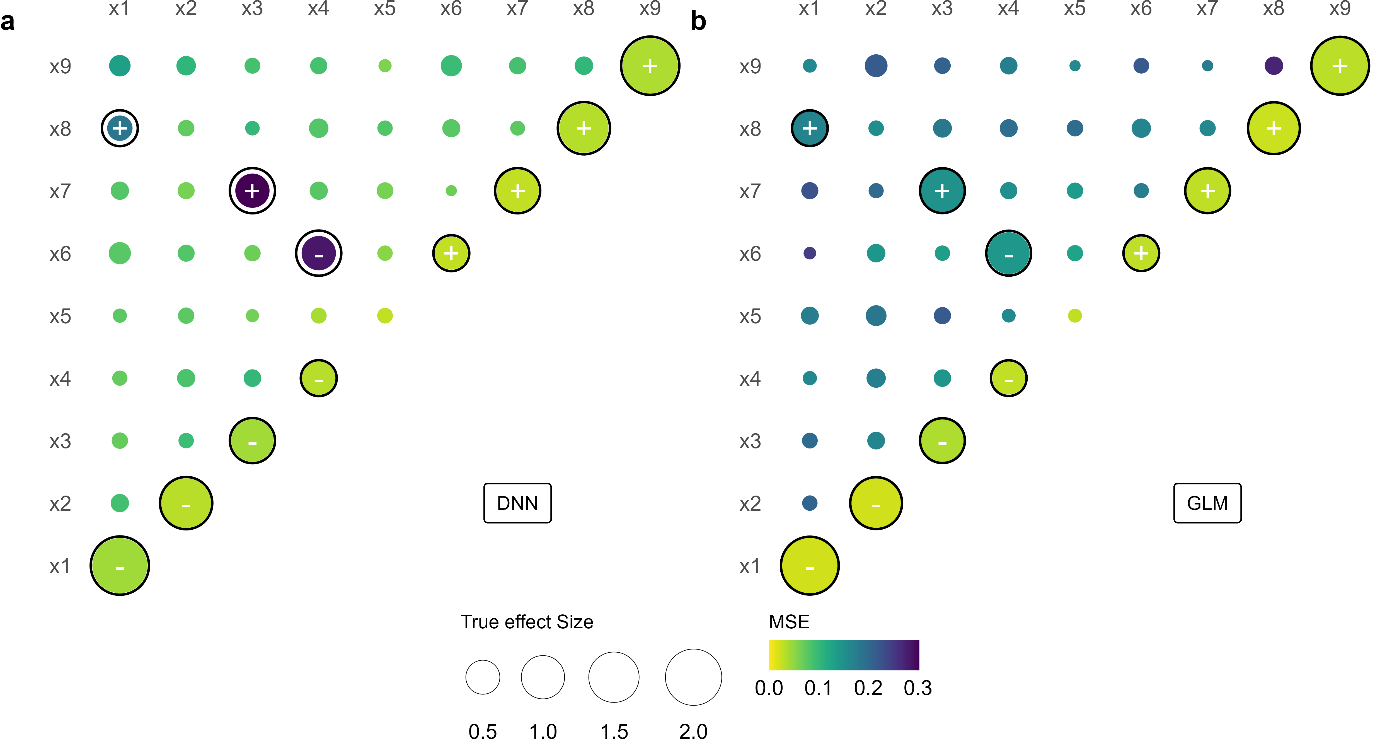
Ce que change un tel réseau est fondamental. Il n’a pas besoin qu’on lui indique à l’avance les interactions à chercher. En ajustant ses paramètres au fil de l’entraînement, il détecte les relations non linéaires entre variables — à condition qu’elles soient présentes dans les données. Là où le modèle classique requiert qu’on formule a priori la bonne interaction, le réseau identifie la structure relationnelle latente (latent relational structure) que les données recèlent.
Mais une difficulté surgit aussitôt : comment rendre ces résultats lisibles pour un écologue ? Un réseau de neurones profond est, par nature, une boîte noire. On sait ce qu’on y entre, on voit ce qui en sort, et l’intérieur ressemble à une ville vue depuis le ciel — des milliers de connexions dont aucune, prise isolément, n’a de sens. C’est ici qu’intervient ce que l’on appelle l’IA explicable (explainable AI, ou xAI) : une couche d’analyse additionnelle qui traduit les sorties du réseau en coefficients interprétables, directement comparables à ceux produits par le modèle classique. Un pont entre la puissance brute du calcul et le langage des biologistes de terrain. Sans ce pont, la méthode resterait un jouet de statisticiens.
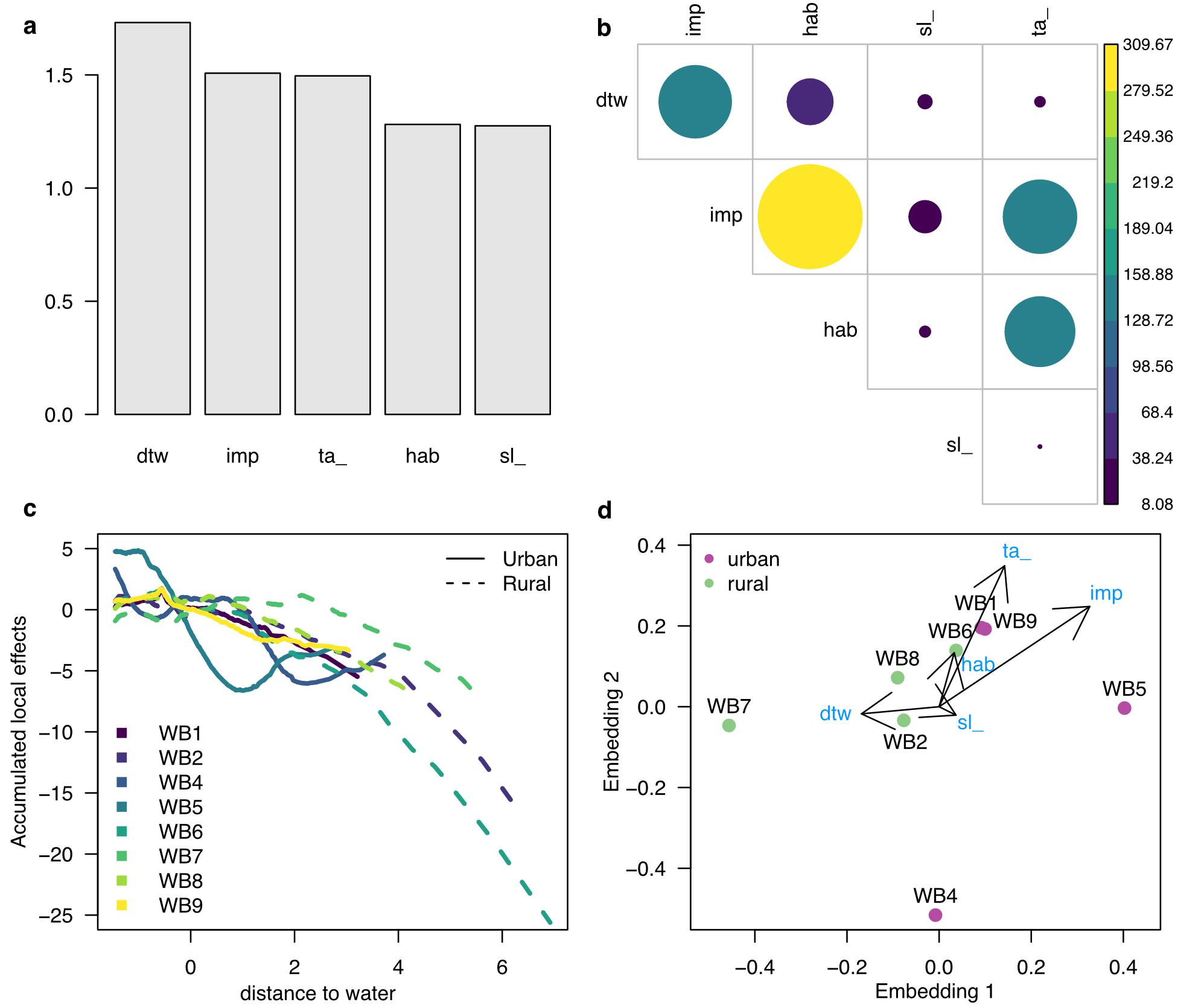
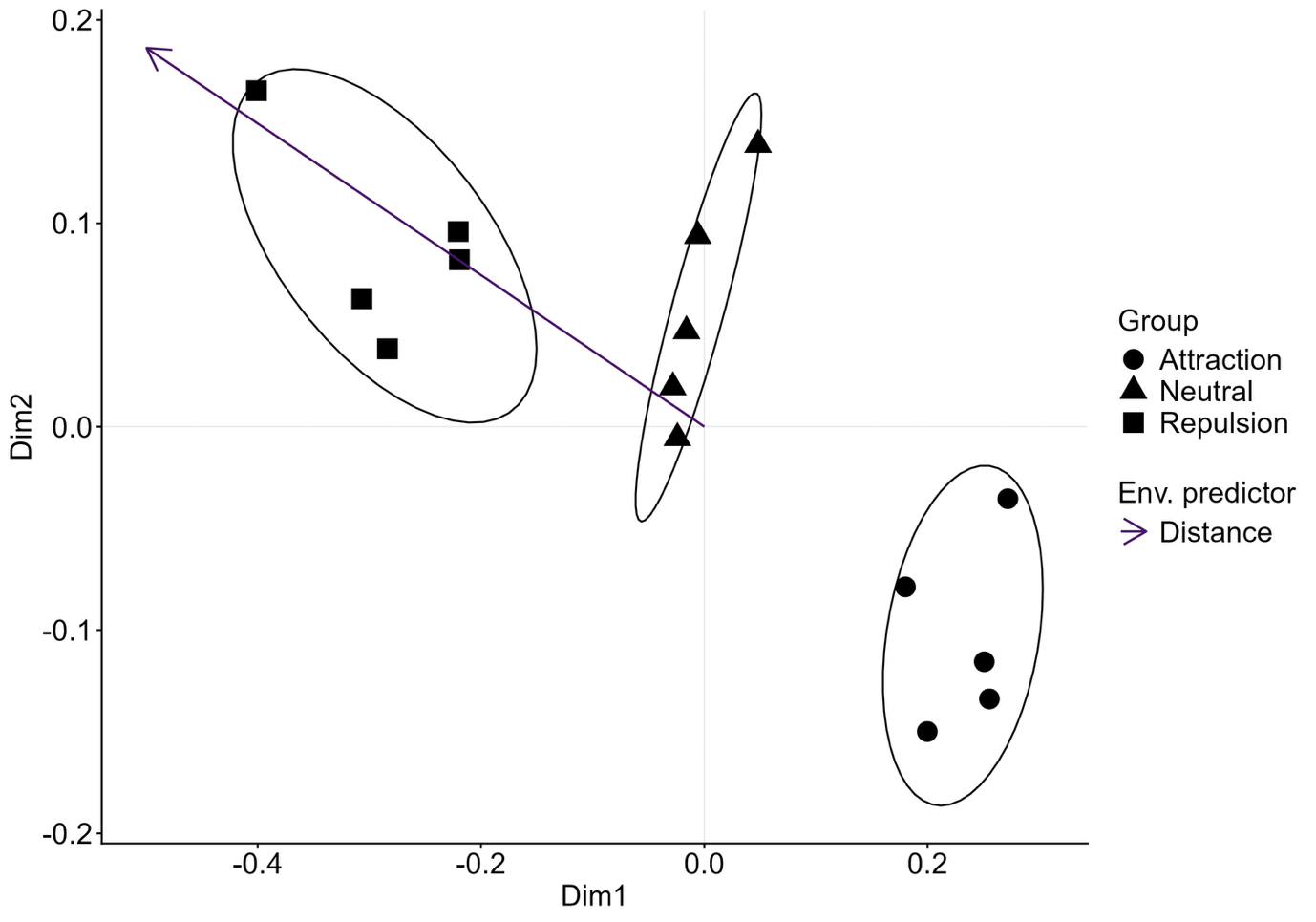
Le package résultant — citoMove, extension directe du amt de Signer — a été mis à l’épreuve sur des données de sangliers équipés de colliers GPS à Berlin. Selon les auteurs, le réseau permettrait de distinguer les individus à profil urbain des individus à profil rural à partir de la seule géométrie de leurs trajets, là où le modèle classique peine à séparer les profils. Ces résultats restent à confirmer sur le texte intégral de la publication, et l’équipe est explicitement prudente : ses conclusions portent sur des cas de test, pas sur une généralisation tous azimuts.
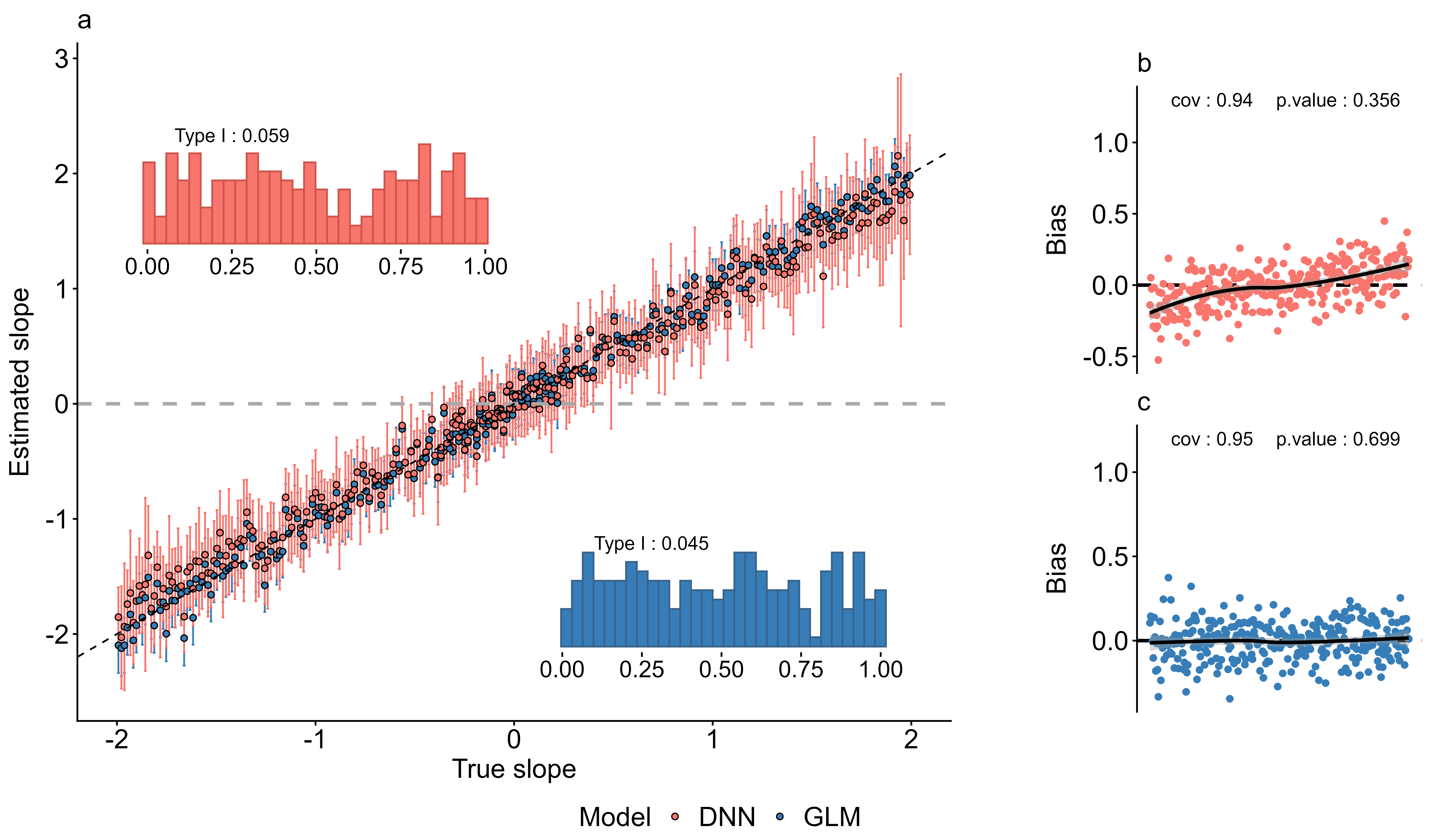
Il y a pourtant une ombre au tableau, et l’équipe ne l’esquive pas. Pour des effets très prononcés, les auteurs rapportent que le réseau tend à légèrement sous-estimer les coefficients — un phénomène qu’ils documentent et discutent dans la publication. Ce n’est pas une anecdote : en écologie du mouvement, sous-estimer un effet fort peut conduire à des conclusions erronées sur les priorités de conservation. La méthode est prometteuse, pas encore infaillible.
Une autre question mérite d’être posée franchement : l’apprentissage profond s’accommode mal des petits jeux de données. Or en éthologie, les suivis GPS coûteux et les populations d’animaux sauvages souvent réduites produisent rarement des millions d’observations. Un réseau de neurones peut très bien ajuster ses paramètres sur du bruit statistique — ce que les spécialistes appellent le surapprentissage (overfitting) — en produisant des résultats apparemment cohérents qui ne reflètent aucune réalité biologique. Les auteurs en sont conscients : citoMove intègre des régularisations adaptées aux petits jeux de données. Mais la prudence reste de mise : la complexité d’un modèle n’est pas une garantie de compréhension.
Ce qui rend cette publication importante, au fond, ce n’est pas seulement la méthode. C’est ce qu’elle annonce. Johannes Signer avait construit amt pour standardiser les pratiques d’une communauté entière d’écologues ; citoMove s’inscrit dans cette continuité, avec l’ambition d’intégrer la puissance des réseaux de neurones sans renoncer à l’interprétabilité que les biologistes exigent à juste titre. L’outil existe, le code est accessible, les données de test sont disponibles.
Il reste à savoir si la communauté s’en emparera — et surtout si elle saura résister à la tentation de croire qu’un algorithme plus complexe livre automatiquement une biologie plus vraie. Un sanglier berlinois à trois heures du matin ne suit pas un modèle. Il suit sa faim, sa mémoire, peut-être une peur ancienne dont nous n’aurons jamais la trace. Les données captent la trace. Le sens, lui, nous échappe encore.
À lire aussi sur Mémorabilité :
- Quand un agent IA prend domicile dans un dépôt Git
- Tâches d’induction, d’analogie et de causalité : des écarts de performance marqués dans les grands modèles de langue
- L’IA ne progresse pas en vase clos : pourquoi son avenir est pluriel et ancré dans les usages sociaux
Figures originales du paper
Sources
- Fronville, Pichler, Signer, Hartig, Kramer-Schadt et al. — citoMove: Deep learning-based step selection functions for animal movement analysis — arXiv:2603.24009
- Package R
amt(Johannes Signer) : référence pour l’analyse de trajectoires animales, dontcitoMoveconstitue une extension directe


