
Chaque seconde, dans un millilitre d’eau de mer, dix millions de virus percent des bactéries. Pas au hasard — avec la précision d’un cambrioleur qui connaîtrait le plan du coffre-fort, l’emplacement de chaque serrure, l’épaisseur de chaque paroi. Les bactériophages, ces prédateurs qui ne s’attaquent qu’aux bactéries, pratiquent l’effraction moléculaire depuis trois milliards d’années. Et depuis trois milliards d’années, on pensait comprendre exactement comment ils s’y prenaient pour sortir de leur cellule hôte une fois la réplication achevée. Trois protéines, trois couches à traverser, trois étapes séquentielles. Un scénario si propre qu’il tenait en un schéma de manuel. Trop propre, peut-être.
Car il y a un détail que personne ne regardait.
Reprenons la chronologie de l’évasion — elle mérite qu’on s’y attarde. Un phage à ADN double brin vient de fabriquer des dizaines de copies de lui-même à l’intérieur d’une bactérie à Gram négatif, disons une Escherichia coli. Le problème : il est enfermé. La cellule est close de toutes parts, protégée par trois enceintes concentriques dont chacune obéit à sa propre logique architecturale.
Premier acte : les holines. Ces protéines s’accumulent dans la membrane interne — la couche la plus profonde — jusqu’à atteindre un seuil critique. Puis, d’un coup, elles y percent des pores. L’image est brutale mais exacte : des charges explosives posées patiemment le long d’un barrage, qui détonent simultanément. La membrane cède. Le contenu du cytoplasme se déverse vers la couche suivante.
Deuxième acte : les endolysines, libérées par les pores des holines, s’attaquent au peptidoglycane. Ce réseau rigide de chaînes de sucres reliées par de courts segments protéiques forme le squelette de la bactérie — sans lui, la pression osmotique interne ferait éclater la cellule comme la peau d’un raisin qu’on presse entre deux doigts. Les endolysines en coupent les mailles une à une. La paroi se fragilise. Chez les bactéries à Gram positif, qui ne possèdent qu’une seule membrane, l’histoire s’arrêterait là : la cellule éclate, les phages s’échappent.
Mais E. coli possède une troisième ligne de défense. La membrane externe — une double couche lipidique supplémentaire, bardée de molécules complexes — enveloppe le tout. Troisième acte, donc : les spanines fusionnent la membrane interne, déjà trouée, avec cette membrane externe, provoquant sa rupture. Le mur d’enceinte s’effondre. Les phages sont libres.
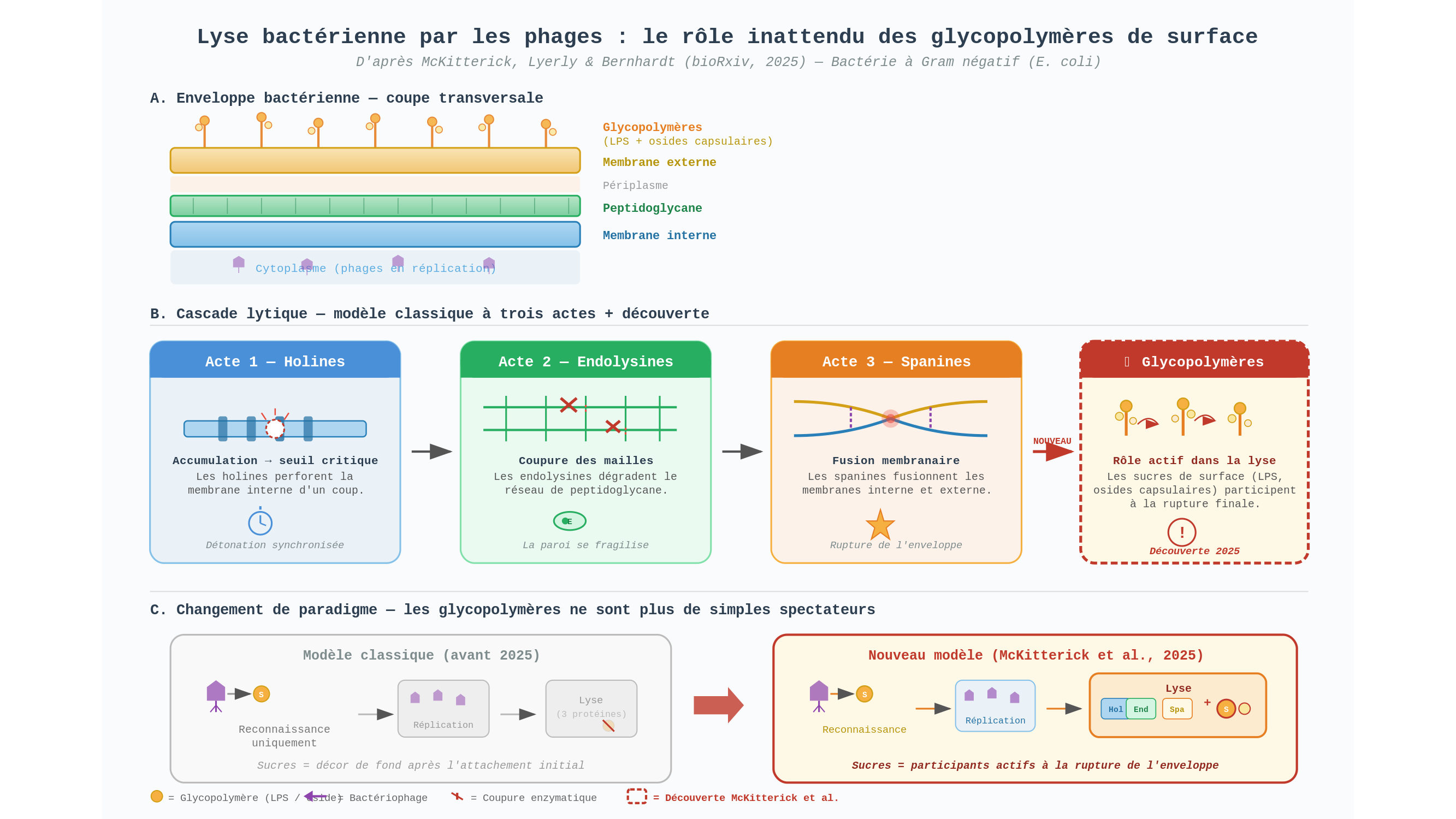
Percer. Éroder. Déchirer. Trois verbes, trois protéines, trois couches. Le scénario semblait complet.
C’est ici qu’Allison McKitterick, Emily Lyerly et Thomas Bernhardt entrent en scène. Dans une prépublication déposée sur bioRxiv en juin 2025, ces trois chercheurs posent une question d’une simplicité presque embarrassante : que fait-on des sucres ?
La surface d’une bactérie à Gram négatif n’est pas nue. Elle est hérissée de glycopolymères — des chaînes de sucres complexes accrochées à la membrane externe. Les lipopolysaccharides (LPS) tapissent la surface d’E. coli comme une pelouse moléculaire dense. Certaines espèces y ajoutent des osides capsulaires, un manteau gélatineux qui enrobe la cellule entière. On connaissait depuis longtemps le rôle de ces sucres au premier acte de l’infection : ce sont eux que le phage identifie pour s’accrocher à sa cible, comme une clé qui trouverait sa serrure dans l’obscurité. Mais une fois le virus entré, on cessait de s’y intéresser. Décor de fond. Figuration muette.
L’équipe de Bernhardt suggère qu’il n’en est rien. Selon leurs travaux, ces sucres de surface interviendraient activement dans les étapes tardives de l’infection — celles de la lyse elle-même. Les glycopolymères ne seraient pas des spectateurs passifs du démantèlement de l’enveloppe, mais des participants fonctionnels dont la présence ou l’absence modifie l’efficacité de la rupture finale.
Imaginez un château médiéval dont les gargouilles, au moment du siège, se détacheraient d’elles-mêmes pour ébranler les remparts. Les ornements travailleraient contre la structure. La forteresse fournirait à l’assaillant ses propres armes.
Le mécanisme précis reste à élucider, et c’est ici que la prudence s’impose. S’agit-il d’un effet mécanique — la déstabilisation structurale de la membrane externe lorsque les sucres sont altérés ? D’une interaction chimique spécifique entre les produits de dégradation des glycopolymères et les spanines ? McKitterick et ses collègues ouvrent la question plus qu’ils ne la referment. Et leur prépublication n’a pas encore été soumise à la révision par les pairs — un filtre que tout lecteur attentif gardera en tête.
Il y a aussi une distinction subtile mais décisive que la biologie cellulaire enseigne à ne jamais négliger : démontrer qu’un composant est impliqué dans un processus n’est pas la même chose que démontrer qu’il y est nécessaire. Le peptidoglycane reste, selon toute vraisemblance, la cible centrale de la lyse. Les glycopolymères pourraient n’en être que les complices circonstanciels — utiles, mais pas indispensables.
Cette réserve faite, les ramifications potentielles dépassent la virologie fondamentale. Depuis une quinzaine d’années, la thérapie phagique — l’utilisation de bactériophages pour traiter des infections résistantes aux antibiotiques — connaît un regain d’intérêt considérable, avec des essais cliniques en cours dans plusieurs pays. Or les bactéries modifient couramment leur profil de glycopolymères pour échapper au système immunitaire. Si ces mêmes sucres jouent un rôle actif dans la lyse, alors une bactérie qui change de manteau pour fuir nos défenses pourrait du même coup altérer sa sensibilité aux phages. La résistance ne se jouerait plus seulement à l’entrée du virus, mais aussi à sa sortie. Le champ de bataille serait plus vaste qu’on ne le pensait.
On peut aller un cran plus loin — en terrain spéculatif. Plusieurs équipes travaillent à isoler des endolysines recombinantes, ces enzymes phagiques produites en laboratoire pour servir directement d’agents antibactériens, sans le phage entier. Si l’efficacité de la lyse dépend en partie de l’environnement glycopolymérique de la bactérie cible, le succès de ces enzymes thérapeutiques pourrait varier d’une souche à l’autre d’une manière que les protocoles actuels ne prennent pas en compte. Un détail moléculaire, des conséquences en cascade.
Pendant des décennies, l’enveloppe bactérienne a été décrite comme un mur — statique, passif, un obstacle que le phage devait franchir par la force. Ce que suggère l’équipe de Bernhardt, c’est que ce mur n’est pas inerte. Ses aspérités, ses sucres greffés, tout ce qui fait d’une bactérie un être chimiquement unique serait pris dans le jeu. Reste la question la plus troublante, celle que trois milliards d’années de coévolution posent sans y répondre : est-ce la forteresse qui trahit, ou le virus qui a retourné chaque pierre de la citadelle contre elle-même ?
Sources
- McKitterick A. C., Lyerly E. W., Bernhardt T. G., ‘Most bacteriophages lyse their host cell to release progeny virions […] spanin-requirement for cell lysis’, bioRxiv prépublication, 2025. DOI: 10.1101/2025.06.24.661397


