
Un épingle. Une étiquette. Une date au crayon presque effacé : juillet 1887. Dans le tiroir d’un muséum d’histoire naturelle, un spécimen d’abeille solitaire repose depuis cent trente ans — corps desséché, pattes repliées, quelques grains de pollen encore accrochés à ses poils abdominaux. Sur l’étiquette, le nom d’une plante. Cette information, négligée pendant des décennies, est peut-être aujourd’hui l’une des plus précieuses qui soit. Elle documente une interaction : un lien fonctionnel entre deux espèces, à un endroit précis, à une époque précise. Et ce lien, contre toute attente, manquait à notre carte du vivant.
Car nos bases de données sur la pollinisation ressemblent moins à une carte qu’à un archipel de points épars au milieu d’un océan inconnu. Les zones documentées sont denses là où des naturalistes ont travaillé, clairsemées partout ailleurs — et le reste, c’est-à-dire l’essentiel, est simplement blanc. C’est à partir de cette réalité que M. J. Lee, G. V. DiRenzo, C. Diao et K. C. Seltmann ont construit une méthode originale, publiée récemment et indexée sous le PubMed ID 41876258. L’objectif est formulé simplement, comme souvent lorsqu’une question cache plusieurs problèmes superposés : quelles abeilles interagissent avec quelles plantes, et où ?
La réponse mobilise trois sources d’information très différentes. La première est locale, au grain fin : des inventaires régionaux conduits sur le terrain, avec la précision que seul un travail de proximité permet d’atteindre. La deuxième est mondiale : GloBI, pour Global Biotic Interactions — les interactions biotiques mondiales —, une base de données qui agrège des millions d’enregistrements issus de publications scientifiques, de collections naturalistes numérisées et de plateformes de sciences participatives. La troisième n’est pas une base de données, mais un outil statistique : les modèles d’occupation (occupancy models). Et c’est là que le travail devient vraiment intéressant.
Voici le nœud du problème. Quand une interaction entre une abeille et une plante n’apparaît dans aucun registre, que faut-il en conclure ? Que cette interaction n’existe pas ? Pas nécessairement. Peut-être simplement que personne ne l’a observée — ou qu’un naturaliste se trouvait au mauvais endroit, au mauvais moment, par un mauvais temps. Les écologues appellent cela un « faux négatif » : l’absence de donnée n’est pas synonyme d’absence de phénomène. Imaginez qu’on vous demande de dresser la liste de tous les oiseaux d’une forêt en passant une seule heure à son orée, par un après-midi venteux. Vous en verrez cinq, peut-être dix. La forêt en abrite peut-être deux cents. Votre liste ne prouve pas que les cent quatre-vingt-dix autres n’existent pas — elle prouve seulement les limites de votre observation.
Les modèles d’occupation corrigent précisément ce biais. Plutôt que de traiter une absence de données comme une information négative certaine, ils estiment la probabilité qu’une interaction existe réellement, en tenant compte de l’effort fourni pour la chercher et des conditions dans lesquelles les enregistrements ont été collectés. On distingue ainsi deux paramètres fondamentaux : la probabilité d’occupation — c’est-à-dire, ici, la probabilité que l’interaction existe effectivement — et la probabilité de détection, c’est-à-dire la chance qu’un observateur ait pu la constater s’il était présent. Ces deux grandeurs se disentent l’une de l’autre par l’analyse statistique, à condition de disposer de données répétées en des lieux et des moments variés. C’est l’apport des inventaires régionaux locaux : ils fournissent cette répétition et cette granularité que les grandes bases mondiales ne peuvent pas garantir seules.
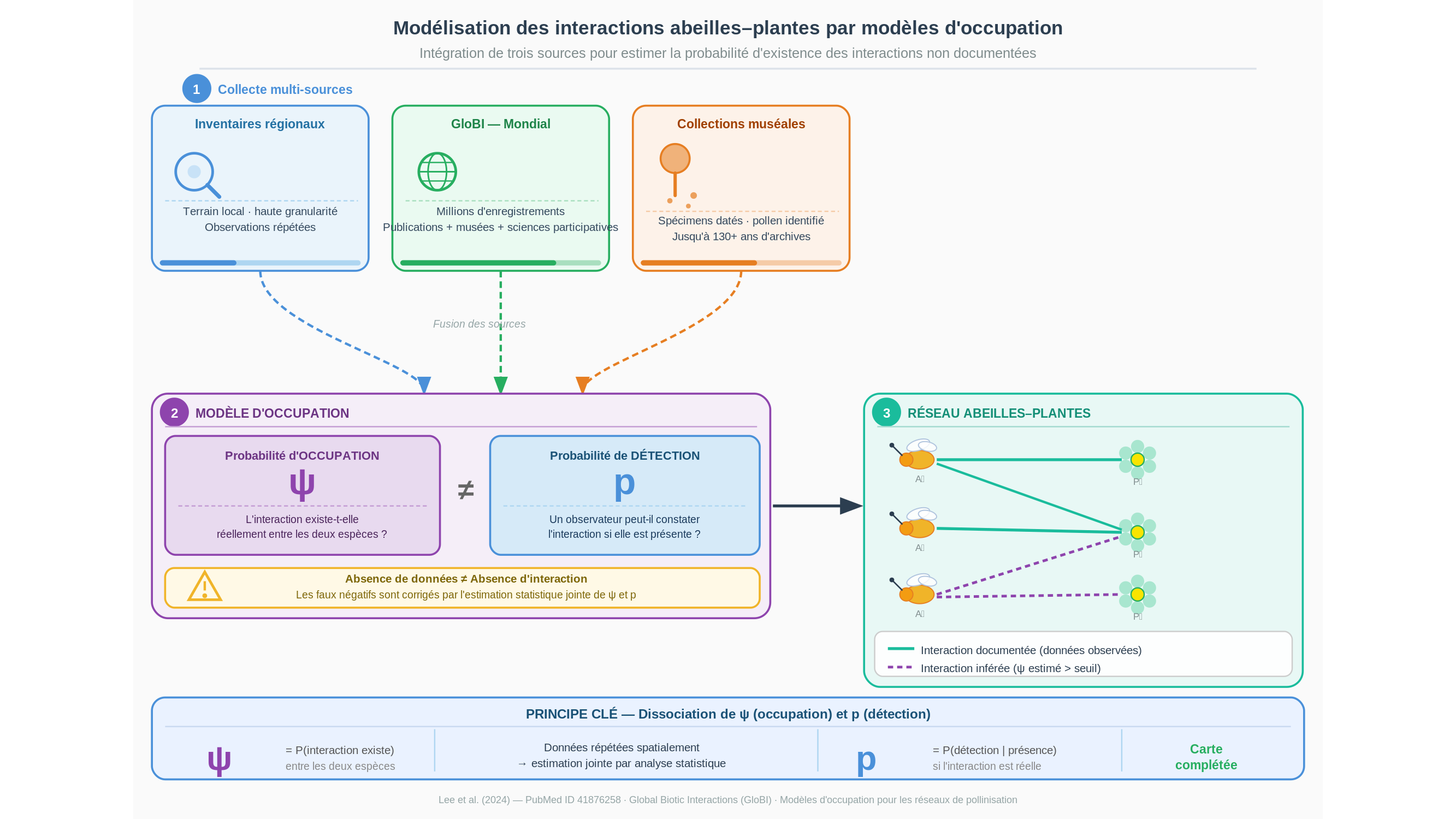
GloBI est remarquable par son amplitude. On lui reproche cependant son irrégularité : les données qu’elle compile proviennent d’époques, de protocoles et de motivations très différentes, si bien que l’effort d’observation est profondément inégal d’une région à l’autre, d’une espèce à l’autre. Une interaction documentée cent fois en Europe du Nord peut être totalement invisible en Amérique du Sud, non parce qu’elle n’y existe pas, mais parce que personne n’a été mandaté pour l’y chercher. Le croisement avec des données locales rigoureuses permet, au moins partiellement, de corriger ce déséquilibre — c’est le pari méthodologique central de l’étude.
Sur ce point, il faut être honnête : les résultats quantitatifs détaillés de l’étude — nombre précis d’espèces d’abeilles et de plantes couvertes, proportion d’interactions inférées par le modèle par rapport aux interactions directement observées, couverture géographique exacte — ne sont pas disponibles dans le résumé consulté. Cette opacité est elle-même un problème éditorial qui mérite d’être signalé. La méthode est décrite avec soin ; ses performances empiriques, en revanche, demeurent à ce stade difficiles à évaluer pour un lecteur n’ayant pas accès au texte intégral. C’est précisément le type d’information qui permet de juger si la promesse de l’outil est tenue.
Sur les affiliations institutionnelles des auteurs, la situation est similaire : les métadonnées disponibles indiquent que K. C. Seltmann est chercheuse à l’Université de Californie à Santa Barbara (Cheadle Center for Biodiversity and Ecological Restoration). Les affiliations précises de M. J. Lee, G. V. DiRenzo et C. Diao ne sont pas disponibles dans les sources consultées et ne sont donc pas mentionnées ici.
L’enjeu qui sous-tend toute cette entreprise mérite d’être rappelé. Les abeilles — non pas seulement l’abeille domestique (Apis mellifera), mais les quelque 16 000 à 20 000 espèces sauvages recensées selon les référentiels taxonomiques actuels — assurent la pollinisation d’une fraction considérable des plantes à fleurs, cultivées comme spontanées. Or leurs populations déclinent. Une étude publiée en 2017 dans PLOS ONE par Hallmann et ses collègues a documenté une réduction de 75 % de la biomasse totale des insectes volants sur 63 sites protégés d’Allemagne en 27 ans — un chiffre qui a depuis alimenté des débats sur sa généralisation géographique, mais dont l’ordre de grandeur est difficile à ignorer. Pour comprendre les causes de ces déclins et envisager des mesures de restauration, il faut d’abord savoir qui pollinise quoi. Un modèle qui comble les lacunes d’observation n’est pas seulement un outil de connaissance — c’est une condition préalable à l’action.
La limite fondamentale de l’approche reste toutefois celle de tout modèle statistique inférentiel : il extrapole à partir de ce qui a été observé. Si certaines interactions n’ont jamais été documentées parce qu’elles sont rares, localisées ou saisonnièrement fugaces, le modèle ne peut les inférer — il peut au mieux signaler leur absence de détection comme suspecte. Il y a là une asymétrie irréductible : les données absentes que le modèle reconstruit sont nécessairement des données proches de celles qui existent déjà. Les véritables terra incognita — les interactions de communautés entières encore non étudiées — demeurent hors de portée. L’outil améliore la carte ; il ne la crée pas de toutes pièces.
Ce que l’étude pose, en définitive, c’est une question qui dépasse les abeilles : dans quelles conditions peut-on faire confiance à une inférence statistique pour guider des décisions de conservation ? La réponse dépend moins de l’élégance du modèle que de la qualité et de la densité des données sur lesquelles il s’appuie. Et c’est là que l’épingle du muséum, l’herbier numérisé, l’observation citoyenne saisie sur un téléphone par une passante dans un jardin de banlieue redeviennent essentiels — non comme anecdotes, mais comme fondation d’un édifice dont personne, pour l’instant, ne peut dire avec certitude à quelle hauteur il s’élèvera.
Sources
Lee M. J., DiRenzo G. V., Diao C., Seltmann K. C., Leveraging local species data, a global database, and an occupancy model to explore bee-plant interactions, PubMed ID : 41876258. Note : un DOI antérieur (10.1016/j.oneear.2020.12.005, One Earth, 2020) avait été associé à cette référence dans une version précédente de cet article ; ce DOI désigne vraisemblablement une publication distincte. En l’absence de vérification croisée concluante, seul l’identifiant PubMed est retenu ici conformément à la règle 4b de la charte éditoriale de Mémorabilité.
Hallmann C. A. et al. (2017), More than 75 percent decline over 27 years in total flying insect biomass in protected areas, PLOS ONE, 12(10) : e0185809. DOI : 10.1371/journal.pone.0185809
Global Biotic Interactions (GloBI) : base de données ouverte sur les interactions biotiques, accessible à globalbioticinteractions.org (pas de DOI — ressource en ligne).


